
60 YEARS OF POMC: From POMC and α-MSH to PAM, molecular oxygen, copper, and vitamin C
- 1Departments of Molecular Biology and Biophysics, University of Connecticut Health Center, Farmington, Connecticut, USA
- 2Department of Neuroscience, University of Connecticut Health Center, Farmington, Connecticut, USA
- Correspondence should be addressed to R E Mains or B A Eipper; Email: mains{at}uchc.edu or eipper{at}uchc.edu
-
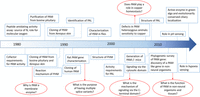 Figure 1
Figure 1Timeline highlighting key developments leading from POMC processing studies to PAM. The landmark study of Bradbury and coworkers provided a means of assaying peptide amidating activity in tissue lysates (Bradbury et al. 1982). Purification, cloning, and structural/mechanistic studies focused on PAM and then expanded to include cell biological studies on secretory granule biogenesis, retrograde signaling from the granule lumen to the nucleus, and the delivery of essential cofactors (ascorbate and copper) to the secretory pathway. Key unanswered questions are marked by red arrows.
-
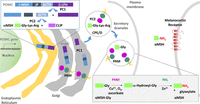 Figure 2
Figure 2POMC processing: amidation of α-MSH. Following the cotranslational removal of its N-terminal signal sequence, POMC moves through the Golgi complex. As luminal pH begins to fall and prohormone convertase 1 (PC1) is activated, the first POMC cleavage produces ACTH biosynthetic intermediate and β-LPH. Subsequent cleavages (upper left box), which occur largely in maturing secretory granules, separate JP from ACTH; the C-terminus of JP can be amidated. Melanotropes, which express both PC1 and PC2, cleave ACTH(1–39) to produce an N-terminal fragment (precursor to α-MSH) and CLIP (corticotropin-like intermediate lobe peptide). The production of α-MSH requires a carboxypeptidase, PAM, and an N-acetyltransferase (not shown). The sequential actions of PHM and PAL on α-MSH-Gly are illustrated (lower right box).
-
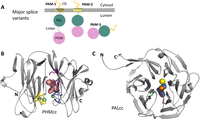 Figure 3
Figure 3Major PAM splice variants and PHM and PAL catalytic core structures. (A) The major splice variants (isoforms) of PAM are shown. Although PAM-1 and PAM-2 are type 1 integral membrane proteins, PAM-3 is a soluble, secreted protein. (B) Crystal structure of rat PHM (PDB identifier: 1OPM) in the oxidized state bound to a substrate, N-acetyl-3,5-diiodotyrosylglycine (shown in pink), rendered here using PyMOL; bound copper, green spheres. The copper-binding site in the N-terminal domain of PHM (CuH, in yellow) is separated from the copper-binding site in the C-terminal domain (CuM, in blue) by an 11 Å solvent-filled cleft; the peptidylglycine substrate and molecular oxygen bind near CuM. Other essential catalytic residues involved in substrate binding (R240, Y318, and M320) are shown in purple. After considering many mechanisms, quantum mechanical tunneling is thought to facilitate electron transfer from the CuH site, through solvent, to the CuM site (Francisco et al. 2004, Klinman 2006, McIntyre et al. 2010). (C). The structure of PAL (PDB identifier: 3FW0) crystallized in the presence of mercury ion (orange) instead of zinc to capture the binding of a nonpeptide substrate (α-hydroxyhippuric acid, in blue) is shown. The six-bladed β-propeller structure of PAL positions Zn near a key Tyr residue (shown in purple) and a key Arg residue (shown in green). The structurally important calcium ion is depicted as a yellow sphere (Chufan et al. 2009).
-
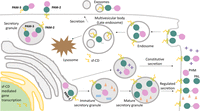 Figure 4
Figure 4PAM trafficking. Membrane PAM (PAM-1, PAM-2) travels through both the biosynthetic and endocytic pathways; soluble PAM (PAM-3) is efficiently packaged into secretory granules and secreted. Cleavage of PAM in the biosynthetic pathway involves the prohormone convertases and cleavages at pairs of basic residues in the linker region between PHM and PAL and immediately following PAL. Cleavages on the cell surface and in the endocytic pathway involve α-secretase and γ-secretase. Phosphorylation of its cytosolic domain affects the ability of PAM to move from the limiting membrane of multivesicular bodies (MVBs) into the intraluminal vesicles. Endocytosed PAM can be returned to the trans-Golgi network, for re-entry into secretory granules, or degraded in lysosomes.
-
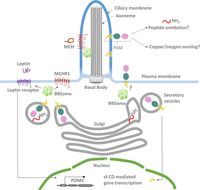 Figure 5
Figure 5Model of PAM function in cilia. PAM localizes to cilia, where it may play roles in peptide amidation and sensing of copper or oxygen. Ciliary PAM may also be subjected to cleavages leading to the release of sf-CD and changes in gene expression. It is not known if the BBSome is essential for PAM trafficking into the ciliary compartment. However, trafficking of the MCHR1 into the cilium requires the BBSome, and leptin receptor localization is affected by loss of BBS components. The leptin/melanocortin system is critical for establishing energy homeostasis. Leptin binding to its receptor leads to upregulation of POMC and satiety responses. It is not known if PAM also plays a role in energy balance in POMC neurons.
- © 2016 Society for Endocrinology










